


 التطور التدريجي للحيوانات والنباتات عبر العصور موضوع مفصل
التطور التدريجي للحيوانات والنباتات عبر العصور موضوع مفصل
التطور:
هو التغير في السمات الوراثية الخاصة بأفراد التجمع الأحيائي عبر الأجيال المتلاحقة. السيرورات التطورية تُحدث تنوعاً حيوياً في كل المستويات التصنيفية، بما فيها الأنواع، أفراد الكائنات الحية، والجزيئات كالدنا والبروتينات.[1]
نشأت الحياة على الأرض ومن ثم تطورت من سلف شامل أخير منذ نحو 3.7 بليون سنة. يُستَدل على الانتواع المتكرر والتخلق التجددي للحياة بالنظر إلى المجموعات المشتركة من السمات المورفولوجية والكيميائية الحيوية، أو من تسلسلات الدنا المشتركة. وهذه السمات والتسلسلات المتماثلة تكون متشابهة أكثر كلما كان السلف المشترك للأنواع أحدث. ويمكن استخدامها في إعادة بناء التاريخ التطوري بالاستناد إلى الأنواع الموجودة حالياً وأيضاً السجلات الأحفورية. أنماط التنوع الحيوي الموجودة تشكلت بفعل الانتواع والانقراض.[2]
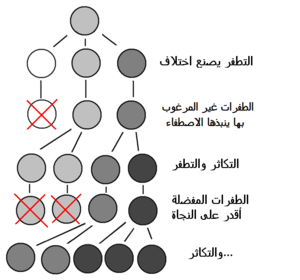
كان تشارلز داروين أول من صاغ محاججة علمية لنظرية التطور الذي يحدث بواسطة الاصطفاء الطبيعي. التطور بواسطة الاصطفاء الطبيعي هو عملية يُستدل عليها من ثلاث حقائق تخص التجمعات: 1- النسل الذي يتم انجابه يكون أكبر عددا من ذاك القادر على النجاة. 2- السمات تتباين بين الأفراد، مما يؤدي لاختلاف معدلات النجاة والتكاثر 3- الاختلافات في السمات هي وراثية.[3] فأفراد التجمع الذين يموتون يُستبدلون بنسل الوالدِين الذين كانوا متكيفين أكثر على النجاة والتكاثر في البيئة التي حدث بها الاصطفاء الطبيعي. وهذه العملية تُنتِج سمات تبدو ملائمة للأدوار الوظيفية التي تقوم بها، وتحافظ عليها.[4] الاصطفاء الطبيعي هو المسبب الوحيد المعروف للتكيف، ولكنه ليس المسبب الوحيد المعروف للتطور. فثمة مسببات غير تكيفية أخرى للتطور، ومنها الطفرات والانحراف الجيني.[5] في مطلع القرن العشرين، تم دمج علم الوراثة مع نظرية داروين للتطور بالاصطفاء الطبيعي في الوراثيات السكانية. وقُبلت أهمية الاصطفاء الطبيعي في توليد التطور في فروع علم الأحياء الأخرى. فضلا عن أن تصورات سابقة حول التطور، مثل نظرية استقامة التطور و"التقدم"، أصبحت مبطلة.[6] ويواصل العلماء دراسة جوانب عدة من التطور عن طريق وضع فرضيات واختبارها، بناء نظريات علمية، استخدام معطيات ملاحظة، واجراء تجارب مخبرية وميدانية. يُجمع علماء الأحياء على أن التحدر مع تغييرات (أي التطور) هو من أكثر الحقائق المؤكدة على نحو موثوق في العلوم.[7] وقد تجاوز تأثير الاكتشافات في علم الاحياء التطوري حدود فروع الأحياء التقليدية، ويتمثل ذلك بتأثيرها الهائل في مجالات أكاديمية أخرى مثل علم النفس التطوري وعلم الإنسان) وفي المجتمع عامةً.
تاريخ الفكر التطوري :
الاعتقاد بأن أنواع الحيوانات يمكن أن تتحدر من أنواع أخرى، يعود إلى زمن الفلاسفة الإغريقيين قبل سقراط مثل أناكسيماندر وإيمبيدوكليس.[10][11] وبخلاف آرائهما المادية، نظر أرسطو إلى كل الأشياء الطبيعية، وليس فقط الكائنات الحية، على أنها تحقيق غير مثالي لاحتمالات طبيعية ثابتة تعرف "بالأشكال" أو "الأفكار" أو (بالترجمة اللاتينية) "الأنواع".[12][13] وهذا كان جزءاً من رؤيته الغائية للطبيعة، إذ كل شيء فيها يؤدي دوراً معداً له في نظام كوني إلهي. اقترح الشاعر والفيلسوف الروماني لوكريتيوس فكرة احتمال حدوث تغيرات تطورية بالكائنات الحية.[14] وشكلت صيغاً مختلفة لهذه الفكرة الفهم العام في العصور الوسطى، وتم ضمها إلى تعاليم المسيحية.[15]
لم يتفق منهج العلم الحديث في القرن السابع عشر مع مأخذ أرسطو، وكان لا بد من البحث عن تفسيرات لظواهر طبيعية بحسب قوانين الطبيعة، والتي كانت تنطبق على كل الأشياء المرئية على حد سواء، ولم تكن هناك حاجة لافتراض أي تصنيفات طبيعية ثابتة أو أي نظام كوني إلهي. ولكن هذا المنهج الجديد أخذ وقتاً ليترسخ في العلوم الأحيائية، والتي أصبحت آخر حصن لمفهوم التصنيفات الطبيعية الثابتة. استعمل جون ري مصطلحاً كان في السابق عاماً أكثر بإشارته إلى التصنيفات الطبيعية الثابتة، ألا وهو "الأنواع"، وذلك للإشارة إلى تصنيفات النبات والحيوان. ولكن بخلاف أرسطو، عرَّف ري بتشدد كل تصنيف من الكائنات الحية على أنه نوع، واقترح أنَّ كل نوع يمكن تمييزه بخواصه التي تدوم مع مرور الأجيال.[16] وكانت هذه الأنواع، وفقاً لري، مصممة من قبل الله، ولكن الاختلافات التي تظهرها سببها الظروف المحلية. التصنيف الحيوي الذي عرضه كارولوس لينيوس عام 1735 كانت الأنواع بحسبه أيضاً ثابتة بموجب تنظيم إلهي.[17]
قام بعض علماء التاريخ الطبيعي الآخرون بنفس الفترة الزمنية بوضع فرضيات عن تغير الأنواع التطوري عبر الزمن وفقاً للقوانين الطبيعية. كتب موبرتيوس عام 1751 عن تغيرات طبيعية تحدث خلال التكاثر وتتراكم عبر أجيال عديدة إلى أن تؤدي لإنتاج أنواع جديدة.[18] واقترح بوفون أنَّ الأنواع قد تتحول إلى كائنات حية مختلفة، واقترح إراسموس داروين أنَّ كل الحيوانات ذات الدم الحار يمكن أن تكون قد تحدرت من كائن حي مجهري واحد (أو "خيط").[19] في عام 1809، قام جان-باتيست لامارك بوضع نظريته "تحول الأنواع"، أولى النظريات العلمية الكاملة للتطور،[20] والتي تصور فيها أنَّ تولداً ذاتياً ينتج باستمرار أشكال حياة بسيطة تطور تعقيدها بسلالات متوازية بنزعة تقدمية فطرية، وأنَّ هذه السلالات، على مستوى محلي، تتكيف مع المحيط عن طريق وراثة التغيرات التي يسببها عدم استعمال الميزات في جيل الآباء.[21][22] (فيما بعد أطلق على هذه السيرورة الأخيرة اللاماركية).[21][23][24][25] استنكر هذه الأفكار علماء تاريخ طبيعي معتمدون واعتبروها فرضية تفتقد للدعم التجريبي. وبالتحديد أصر جورج كوفييه على أنَّ الأنواع ثابتة ولا تربطهم صلة قرابة، والتشابهات بينهم تعكس تصميمها الإلهي الخادم للحاجات الوظيفية. وفي هذه الأثناء كان ويليام بيلي قد طوَّر أفكار ري عن التصميم الهادف فأصبحت لاهوتاً طبيعياً، والذي اقترح أنَّ التكيفات المعقدة دليلٌ على التصميم الإلهي، وقد أعجب تشارلز داروين بها.[26][27][28]
إنَّ الإنطلاق الرئيسي من مفهوم الأنواع الثابتة في الأحياء بدأ مع نظرية التطور عن طريق الاصطفاء الطبيعي، والتي صاغها تشارلز داروين. ذكر داروين أنَّ نمو الكثافة السكانية قد يؤدي إلى "صراع من أجل البقاء"، بحيث تسود التنوعات المحبذة فيما البقية تتلاشى. مع كل جيل، الكثير من النسل لا يقدر على البقاء حتى سن البلوغ والتكاثر بسبب الموارد المحدودة.[30][31][32][33] عمل داروين على تطوير نظرية "الاصطفاء الطبيعي" الخاصة به منذ عام 1838، حتى أرسل له ألفرد راسل والاس نظريته المشابهة عام 1858. وقام كل منهما على حده بعرض دراسته على جمعية لندن اللينية (بالإنجليزية).[34] في آخر عام 1859، الطريقة التي شُرح بها الاصطفاء الطبيعي في كتاب أصل الأنواع لداروين أدت لقبول واسع آخذ بالازدياد للتطور الدارويني. وقام توماس هنري هكسلي بتطبيق أفكار داروين على البشر، وذلك بالاستناد إلى علم الأحياء القديمة والتشريح المقارن من أجل تقديم دليل قوي على أن البشر والقردة لديهم سلف مشترك. والبعض انزعج من هذه الفكرة لكونها تتضمن تلميحاً إلى أنه ليس للبشر مكانة مميزة في الكون.[35]
هناك أمور بقيت لغزاً مثل الآليات المحددة التي تحدث بها الوراثة التناسلية ومصدر السمات الجديدة. ولذلك قام داروين بتطوير نظريته المؤقتة المسماة شمولية التخلق.[36] في عام 1865، أعلن غريغور مندل أنَّ السمات تُورث بشكل يمكن التنبؤ به بواسطة التوزيع المستقل وفصل العوامل (أصبحت تعرف بالجينات فيما بعد). في نهاية المطاف استبدلت قوانين مندل الوراثية معظم نظرية شمولية التخلق لداروين.[37] قام أوغست وايزمان بالتمييز بين الخلايا المُنتِشة (البيوض والحيوانات المنوية) وبين الخلايا الجسدية، بحيث وضَّح أنَّ الوراثة تمرُّ عن طريق الخط الإنتاشي فقط. هيوغو دا فريس ربط نظرية شمولية التخلق لداروين مع تمييز وايزمان بين الخلية الجسدية والإنتاشية، واقترح أنَّ الجينات الشاملة في نظرية داروين كانت متركزة في نواة الخلية، وعندما يُعَبَّر عنها جينياً تنتقل إلى السيتوبلازما لتقوم بتغيير تركيبات الخلايا. دا فريس كان أيضاً أحد الباحثين الذين جعلوا أعمال مندل تشتهر، إذ اعتقد أنَّ السمات المندلية توافقت مع انتقال الاختلافات الوراثية عبر الخط الإنتاشي.[38] من أجل تفسير كيفية نشوء الاختلافات، طوَّر دا فريس نظرية طفور أدت إلى ظهور انشقاق مؤقت بين الذين قبلوا التطور الدارويني وبين الأخصائيين في الإحصاء الحيوي الذين تحالفوا مع دا فريس.[22][39][40] وفي مطلع القرن العشرين، قام روادٌ في مجال الوراثيات السكانية، مثل جون هالدين، سيوال رايت، ورونالد فيشر، بوضع أسس التطور في فلسفة إحصائية. وهكذا تم حل التناقض الخاطيء بين نظرية داروين، الطفرات الجينية، والوراثة المندلية.[41]
في سنوات العشرينات والثلاثينات، ربط الاصطناع التطوري الحديث بين الاصطفاء الطبيعي ونظرية الطفور والوراثة المندلية، ووحَّدها في نظرية تنطبق بوجه عام على أي فرع في علم الأحياء. من خلال الاصطناع التطوري الحديث يمكن تفسير الأنماط الملاحظة في الأنواع بالتجمعات الأحيائية، وذلك باستعمال الأحافير الانتقالية في علم الأحياء القديمة، وأيضاً بواسطة الآليات الخلوية المعقدة في علم الأحياء النمائي.[22][42] إنَّ نشر مبنى الدنا لجيمس واتسون وفرنسيس كريك عام 1953 عرض أساساً مادياً للوراثة.[43] وقد حسَّن علم الأحياء الجزيئي من فهمنا للعلاقة بين النمط الظاهري والنمط الجيني. حدثت تقدمات في النظاميات الفيلوجينية، وبتخطيط انتقالات السمات في إطار مقارن وقابل للاختبار باستعمال الشجرة التطورية.[44][45] في عام 1973، كتب عالم الأحياء التطوري ثيودوسيوس دوبجانسكي: "لاشيء في علم الأحياء يكون منطقياً إلا في ظل التطور"، وذلك لأن التطور وضَّح العلاقات بين حقائق ظهرت على أنها منفصلة في التاريخ الطبيعي، وجعلها مجموعة معارف تفسيرية متماسكة تصف وتتنبأ بالكثير من الحقائق الملاحظة عن الحياة في هذا الكوكب.[46]
ومنذ ذلك الحين، تم توسيع الاصطناع الحديث أكثر ليشمل تفسير الظواهر الأحيائية في السلم الكامل والتكاملي للتدرج الحيوي، من الجينات للأنواع. وسُمي هذا التوسيع "بإيكو-إيفو-ديفو" (بالإنكليزية: eco-evo-devo).[47][48][49]
الوراثة
التطور في الكائنات الحية يحدث بفعل التغيّرات التي تطرأ على السمات الوراثية، وهي صفات محددة قابلة للتوريث في الكائن الحي. فمثلاً، لون العينين في الإنسان يُعَدّ صفة وراثية، إذ يمكن للشخص أن يرث سمة "العينين البنيتين" من أحد أبويه.[50] الجينات هي التي تتحكم بهذه السمات الوراثية. مجموعة الجينات الكاملة الموجودة في الجينوم الخاص بالكائن الحي تُسَمَّى بالنمط الجيني لذاك الكائن.[51]
مجموعة السمات الظاهرية التي تشكل بنية الكائن الحي وسلوكه تُسَمَّى النمط الظاهري لذاك الكائن. هذه السمات تتأتى عن تفاعل نمط الكائن الحي الجيني مع بيئته،[52] ولهذا السبب الكثير من السمات الظاهرية لا تُوَرَّث. على سبيل المثال، االتسفع يحدث نتيجة لتفاعل النمط الجيني للشخص مع أشعة الشمس، ولذلك الذرية لا ترث السمرة. إلا أنَّ الاختلاف بين الأفراد بالنمط الجيني ينتج عنه تفاوت بقابلية التأثر بأشعة الشمس. أبرز مثال على ذلك هم من يملكون سمة المهق (الألبينية)، إذ لا يمكنهم الاسمرار أبداً وهم أكثر عرضة للإصابة بحروق الشمس.[53]
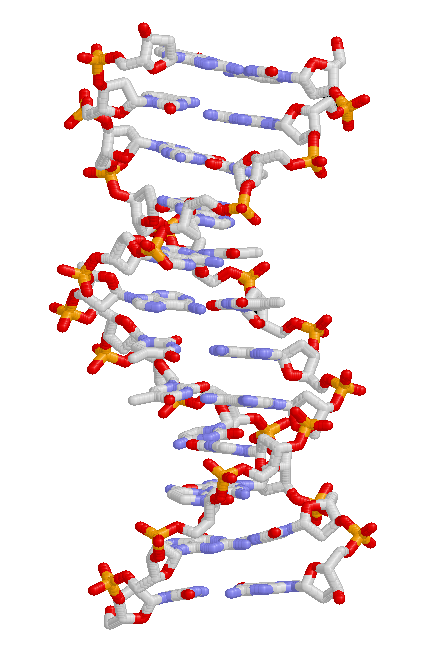
تُنقل السمات الوراثية من جيل لآخر بواسطة الدنا، وهو جزيء يحتوي على معلومات وراثية.[51] وهو بوليمر طويل مُكوَّن من أربعة أنواع من النوكليوتيدات. إنَّ تسلسل هذه النيوكليوتيدات في جزيء الدنا هو الذي يحدد المعلومات الوراثية، مثلما يحدد تسلسل الأحرف معنى الجملة. قبل كل انقسام خلوي، يتضاعف جزيء الدنا الخاص بالخلية، فتحصل كل من الخليتين الابنتين على سلسلة دنا تحمل نفس المعلومات الوراثية. كل قسم من الدنا يمثِّل وحدة وظيفية واحدة يسمَّى بالجين؛ الجينات المختلفة تحتوي على تسلسلات مختلفة من النيوكليوتيدات. تنتظم تسلسلات الدنا في الخلايا لتشكل بُنى مكَدَّسة تُعْرَف بالكروموسومات. الموضع المحدد الذي يحتله تسلسل دنا معين في الكروموسوم يُسمَّى الموضع الكروموسومي (locus). تسلسلات الدنا التي تشغل مواضع كروموسومية متناظرة قد تختلف بين الأفراد بترتيب النيوكليوتيدات، وهذه الصيغ المختلفة لتسلسلات الدنا يُطلق عليها ألائل. الاختلاف بين تسلسلات الدنا يمكن أن يحدث بسبب الطفرات، وينتج عن ذلك ألائل جديدة. عندما تحدث طفرة في الجين، فإنَّ الأليل الجديد الناتج يمكن أن يؤثر على السمات التي يتحكم بها ذاك الجين، مما يؤدي بالتالي لحدوث تغيير في النمط الظاهري للكائن الحي الذي حدثت به الطفرة.[54] إلا أنَّ هذه العلاقة البسيطة المباشرة بين الأليل والنمط الظاهري تتحقق في بعض الحالات فقط. فمعظم سمات النمط الظاهري هي أكثر تعقيداً من ذلك، وتكون ناتجة عن تفاعل بين عدة جينات (بالإنجليزية) تتحكم بهذه السمات.[55][56]
لقد أثبتت نتائج دراسات حديثة أنَّ بعض التغيّرات القابلة للتوريث لا يمكن أن تعزى لتغيّر تسلسل القواعد في الدنا. هذه الظاهرة تُصَنَّف على أنها نظام وراثي لاجيني.[57] وقد تم اكتشاف أنظمة وراثية لاجينية على مستوى الكائنات الحية في المجالات التالية:
- دمغ الكروماتين عن طريق مثيلة الدنا (بالإنكليزية: DNA methylation).
- عمليات أيضية مستديمة ذاتياً (بالإنكليزية: self-sustaining metabolic loops). تقوم فيها البروتينات الناتجة عن جين ما بإطالة فعاليَّة نفس الجين.
- إسكات الجين عن طريق التداخل الرناوي (بالإنجليزية).
- بنية البروتينات الفراغية ثلاثية الأبعاد، مثل البريون.[58][59]
اقترح العلماء المختصون بعلم الأحياء النمائي أنَّ التفاعلات المعقدة التي تحدث في الشبكات الجينية وعمليات التواصل بين الخلايا، قد تتسبب في نشوء تنوعات جينية قابلة للتوريث يمكن أن تمثِّل الأساس الذي تقوم عليه بعض الآليات في التكيفية النمائية والمتانة. [60] التوريث قد يحدث أيضاً على نطاقات أوسع، فمثلاً التوريث البيئي الذي يحدث كنتيجة لعمليات بناء الموطن (بالإنكليزية: niche construction)، يعرَّف على أنه النشاطات المنتظمة والمتكررة التي تقوم بها الكائنات في بيئتها. وهذا يولد، بمرور الزمن، إرثاً من التأثيرات البيئية التي تعدل في نظام الانتخاب الطبيعي الذي سيقع على الأجيال اللاحقة. هكذا ترث الذرية الجينات، بالإضافة للخصائص البيئية التي تولدت عن نشاطات الأسلاف البيئية.[61] هنالك أمثلة أخرى على وراثات لا تتحكم بها الجينات بشكل مباشر، ومنها توريث السمات الثقافية، وأيضاً ما يعرف بالنشوء التعايشي.[62][63]
التنوع
النمط الظاهري لأي كائن حي يتأتى من نمطه الجيني وتأثره بالبيئة التي يعيش بها. جزء كبير من الاختلاف في الأنماط الظاهرية بين الكائنات الحية يعود للاختلاف في أنماطها الجينية.[56] الاصطناع التطوري الحديث يُعرّف التطور على أنه التغير الذي يطرأ على التنوع الجيني (بالإنكليزية: genetic variation) بمرور الزمن. يمكن قياس مقدار التنوع الجيني في التجمعات من خلال حساب تواتر الألائل. تواتر أليل معين هو نسبة عدد نسخ هذا الأليل إلى عدد نسخ كل ألائل نفس الجين، ومدى تواتر الأليل قد يزداد أو يقل بالمقارنة مع الألائل الأخرى لنفس الجين. التنوع يختفي عندما يصل أليل جديد لمرحلة التثبيت (بالإنكليزية: fixation)، هذا يعني إما أنه يختفي تماماً أو يستبدل الأليل السلفي بالكامل.[64]
الانتخاب الطبيعي يسبِّب التطور فقط إذا توفَّر لدى التجمع قدرٌ كافٍ من التنوع الجيني. قبل اكتشاف علم الوراثة المندلي، إحدى الفرضيات السائدة كانت الوراثة الخلطية. ولكن بالوراثة الخلطية التنوع الجيني يُفقد بسرعة، الأمر الذي يجعل التطور من خلال الانتخاب الطبيعي بعيد الاحتمال. مبدأ هاردي-واينبيرغ يقدم حلاً للكيفية التي يتم بها الحفاظ على التنوع في التجمع الأحيائي بتوافق مع الوراثة المندلية. تواتر الألائل في التجمع الأحيائي يبقى ثابتاً بغياب كلٍ من الاصطفاء، الطفرات، الهجرة، والانحراف الوراثي.[65]
مصادر التنوع الجيني هي: حدوث الطفرات في المادة الوراثية، وإعادة خلط الجينات عن طريق التكاثر الجنسي أو بواسطة الهجرة ما بين التجمعات الأحيائية (ما يُعْرَف بانسياب الجينات). رغم تَسَبُّب الطفرات وانسياب الجينات بإدخال مستمر للتنوعات الجينية، إلا أنَّ الأفراد التابعين لنفس النوع يتطابقون في معظم الجينوم الخاص بهم.[66] ولكن حتى الاختلافات الضئيلة نسبياً في النمط الجيني قد تؤدي لاختلافات جذرية في الأنماط الظاهرية. فالاختلاف بين جينوم الإنسان وجينوم الشامبانزي، على سبيل المثال، هو حوالي 5% فقط.[67]
الطفرة:
الطفرات هي تغيّرات تحدث في تسلسل الدنا الخاص بجينوم خلية ما. عندما تحدث الطفرات، قد لا يكون لها أي تأثير، أو قد تُحدث تغييراً في النواتج الجينية (بالإنجليزية)، أو قد تعطل الجين وتمنعه عن العمل. وبناءً على دراسات تمَّت على ذباب الفاكهة (بالإنكليزية: Drosophila melanogaster)، أشير إلى أنَّ التغيير الذي تُحدِثه الطفرة في بروتين أنتجه الجين، من الممكن أن يكون مؤذياً، إذ تصل نسبة الطفرات ذات التأثيرات الضارّة إلى 70%، أما البقية منها فتكون إما محايدة، أو نافعة لدرجة ضئيلة.[68]
تحدث الطفرات أحياناً عن طريق تضاعف أجزاء كبيرة من الكروموسوم (عادةً بواسطة التأشيب الجيني)، الأمر الذي قد يُدخل إلى الجينوم نسخاً زائدة من الجين.[69] وتمثِّل هذه النسخ الزائدة المصدر الرئيسي للمواد الأولية اللازمة لنشوء جينات جديدة.[70] هذا الأمر مهم لأن معظم الجينات الجديدة تتطور ضمن عائلات جينية من جينات سابقة الوجود ترجع لأسلاف مشتركة.[71] فمثلاً، العين البشرية تستعمل أربعة جينات لتصنع التراكيب التي تستشعر الضوء: ثلاثة لرؤية الألوان، وواحد للرؤية الليلية. وجميعها تنحدر من سلف مشترك واحد.[72]
يمكن أن تتولد جينات جديدة من جين سلفي عندما تحدث طفرة في نسخة زائدة له ومن ثم تكتسب وظيفة جديدة. وهذه السيرورة تصبح أسهل ما إن يتضاعف الجين، وذلك لأن إضافية النظام (بالإنكليزية: redundancy) تزداد، إذ تكتسب إحدى النسختين وظيفة جديدة بينما تحافظ النسخة السلفية على وظيفتها الأصلية.[73][74] وهناك أنواع أخرى من الطفرات يمكنها أن تُوَلِّد جينات جديدة كلياً من أجزاء الدنا غير المُشَفِّرَة (بالإنكليزية: noncoding DNA). [75][76] توليد جينات جديدة يمكن أن يتم أيضاً عن طريق تضاعف قطع صغيرة من جينات مختلفة، بحيث تختلط هذه القطع فيما بعد، وتتحِّد لتكوِّن جينات جديدة ذات وظائف جديدة.[77][78] في هذه الحالة، أي عندما تُجمَّع جينات جديدة عن طريق خلط قطع موجودة سابقاً، فإنَّ النطاقات البروتينية (بالإنجليزية) الناتجة عن ترجمة هذه الجينات تتصرف كوحدات ذات وظائف مستقلة وبسيطة، التي يمكن تجميعها لإنشاء توليفات جديدة بوظائف جديدة ومعقدة.[79] يُعَدّ مثالاً على ذلك الإنزيم المُصنِّع لمتعدد الكيتيد (بالإنكليزية: polyketide synthase) - إنزيم كبير مسؤول عن تصنيع المضادات الحيوية - الذي يحتوي على ما يقارب المئة نطاق بروتيني مستقل. وكل نطاق يقوم بخطوة مخصصة له من العملية الكلية، وبذلك يسّرع ويحفّز الإنتاج، على غرار نظام خط الإنتاج
الجنس والتأشيب
في التكاثر اللاجنسي، تُوَرَّث الجينات سوياً، بحيث تكون مترابطة. فهي لا تختلط مع أي جينات لكائنات حية أخرى عند التكاثر. بينما في التكاثر الجنسي، يرث النسل مزيجاً عشوائياً من كروموسوما الأبوين، الذي ينشأ بموجب مبدأ التوزيع المستقل (بالإنكليزية: independent assortment)، الذي ينصّ على أنَّ توارث صفة معينة بنمطٍ مندلي لا يؤثر على توارث سمات أخرى بأي شكلٍ من الأشكال. وفي سيرورة أخرى ذات صلة، المعروفة بالتأشيب المتماثل، يتمّ في الكائنات الحية الجنسية تبادُل الدنا بين كروموسومين متماثلين.[81] لا ينتج عن التأشيب أو إعادة التوزيع أي تغيير في تواتر الألائل، وإنما في الترابطات بينها، مما ينتج سلالة ذات توليفات جديدة من الألائل.[82] العملية الجنسية في العادة تزيد من التنوع الجيني، وفي بعض الأحيان تزيد من معدل التطور.
انسياب الجينات
الانسياب الجيني (بالإنكليزية: gene flow) هو تبادل الجينات الذي يتم بين التجمعات الأحيائية أو الأنواع،[85] وبذلك يمكن أن يمثِّل الانسياب مصدراً لتنوعات جديدة بالنسبة للتجمع أو النوع. ما يمكن أن يسبب الانسياب الجيني هو تنقل الأفراد بين تجمعات أحيائية منفصلة. فالانسياب قد يسببه، على سبيل المثال، تنقل الفئران بين المناطق الداخلية والساحلية. أو، كمثال آخر، تنقل اللواقح بين تجمعات الأعشاب المُتحمِّلة للفلزات الثقيلة وتلك الحساسة لها.
انتقال الجينات بين الأنواع يحدث من خلال تهجين الأحياء، أو نقل الجينات الأفقي. إنَّ ما يُقصد بهذا الأخير هو نقل المعلومات الوراثية من كائن حي لآخر ليس من ذريته. هذه العملية هي شائعة أكثر ما يكون لدى البكتيريا.[86] وفي الطب، هذا الأمر يسهم في نشر مقاومة المضادات الحيوية. فحين تحصل إحدى البكتيرات على الجينات التي تُكْسِبُها المقاومة، يصبح بإمكانها نشرها بسرعة لأنواع أخرى من البكتيرات.[87] في بعض الحالات قد تنتقل الجينات من بكتيريا إلى الخلايا حقيقيات النواة. وقد أثبت هذا مخبريا في حالات كخميرة الجعة (بالإنكليزية: Saccharomyces cerevisiae) وخنافس لوبيا أزوكي (بالإنكليزية: Callosobruchus chinensis). [88][89] وأحد الأمثلة على الانتقالات الجينية واسعة النطاق هو الدوارات العلقية (بالإنجليزية)، إذ أنها تلقَّت جينات من البكتيرات، الفطريات والنباتات.[90] الفيروسات أيضاً يمكنها نقل معلومات وراثية بين الكائنات، متيحةً للجينات الانتقال حتى عبر الامبراطوريات.[91] وقد حدث انتقال جيني على نطاق واسع أيضاً بين أسلاف الخلايا حقيقية النواة وأسلاف البكتيرات، وذلك خلال الفترة التي تمَّ فيها اكتساب الصانعات اليخضورية، والمايتوكوندريا. ومن المحتمل أنَّ حقيقيات النواة كانت قد نشأت من الانتقال الأفقي للجينات فيما بين البكتيرات والعتائق.
آليات التطور:
من منظور الدارونية الجديدة (بالإنكليزية: Neo-Darwinism)، التطور يحدث عندما يتغير تواتر الأليل في تجمع أحيائي يتزاوج أفراده فيما بينهم،[65] كما في حال ازدياد تواتر أليل اللون الأسود في تجمع من العثات. الآليات التي تؤدي لتغيّر توافر الأليل تتضمَّن: الاصطفاء الطبيعي، الانحراف الوراثي، الترافق الجيني، الطفرات، وانسياب الجينات.
الاصطفاء الطبيعي
التطور من خلال الاصطفاء الطبيعي هو العملية التي تصبح فيها الطفرات الجينية التي تحسن من فرص تكاثر النوع شائعة أكثر في الأجيال التجمع اللاحقة. كثيراً ما توصف هذه الآلية بأنها بديهية، وذلك لأنها تنبع من ثلاث حقائق بسيطة:
- يوجد تنوع قابل للتوريث في كل تجمُّع أحيائي.
- عدد النسل الكلّي الذي تنجبه الكائنات الحية يزيد عن عدد النسل الذي يستطيع النجاة والتكاثر.
- يتباين أفراد هذا النسل بمقدرتهم على النجاة والتكاثر.
تخلق هذه الظروف تنافساً على فرص البقاء والتكاثر بين الأفراد. وعلى هذا، الكائنات الحية التي تحمل سمات تعطيها أفضلية على منافسيها، تعيش وتتكاثر لتُوَرِّث تلك السمات النافعة للأجيال اللاحقة، في حين لا تُوَرَّث السمات التي لا تمنح أي أفضلية لأصحابها.[93]
الفكرة المحورية في الاصطفاء الطبيعي هي الصلاحية التطورية (بالإنكليزية: evolutionary fitness) للكائن الحي.[94] الصلاحية تُقاس بقدرة الكائن الحي على البقاء والتكاثر، وهذا ما يُحدد حجم مساهمته بتوريث الجينات للأجيال اللاحقة.[94] على أن صلاحية الكائن لا يُعَبَّر عنها بعدد النسل الذي ينجبه، وإنما بنسبة الأفراد الحاملين لجيناته في الأجيال اللاحقة.[95] على سبيل المثال، الكائن الحي القادر على النجاة بفعالية والتكاثر بسرعة، لكن بنفس الوقت ذريته ضعيفة وغير قادرة على النجاة والتكاثر، تُعتبر مساهمته الجينية للأجيال اللاحقة قليلة، مما يجعله قليل الصلاحية.[94] عندما يزيد أحد الألائل الصلاحية أكثر من غيره، فإنَّه سيزداد توافراً في التجمُّع مع ظهور كل جيل جديد. فالانتخاب "يحابي" هذا الأليل. البقاء المُعَزَّز والخصوبة الزائدة هي من الأمثلة على سمات تزيد من الصلاحية. فيما الأليل الضارّ أو الأقل فائدة يحطّ من الصلاحية، وبالتالي يقلّ توافره في التجمع. أي أن الانتخاب "ينبذ" هذا الأليل.[96] تجدر الإشارة إلى أن صلاحية الأليل ليست معطًى ثابتاً، إذ مع تغيّر الظروف البيئية، السمات الضارّة أو المحايدة سابقاً قد تصبح مفيدة. وبنفس الطريقة، السمات المفيدة سابقاً قد تصبح ضارّة.[54] أي أنَّ مسار الاصطفاء بهذه الطريقة يتم باتجاه عكسي. لكن مع ذلك، التطور لا يمكن أن يكون عكوسياً أيضاً؛ فالسمات التي فُقِدَت فيما مضى، لا يمكن أن تظهر ثانيةً باتباع نفس المسار التطوري (أنظر مبدأ اللاعكوسية لدولو (بالإنجليزية)).
الاصطفاء الطبيعي الواقع على سمات كميَّة ذات قيم متباينة، كالطول مثلاً، يُصَنَّف لثلاثة أنواع. الأول هو الاصطفاء الاتجاهي (بالإنكليزية: directional selection). وهو يحابي قيمة معينة للسمة، مما يزيد شيوعها لتصبح هي القيمة المتوسطة، فتتحرك دالة السمة باتجاه اليمين أو اليسار على مرّ الزمن، كما في حال ازدياد طول أفراد نوع ما بمعدّل بطيء.[99] النوع الثاني، الاصطفاء التمزقي (بالإنكليزية: disruptive selection)، يحابي قيمتان حرجتان للسمة، وينبذ القيمة المتوسطة. فيصبح للسمة منوالان. هذا يحدث، على سبيل المثال، عندما تكون الأفضلية في البيئة إما لكائنات طويلة أو قصيرة، ولكن ليس لمتوسطة الطول. النوع الثالث والأخير هو الاصطفاء التثبيتي (بالإنكليزية: stabilizing selection). فيه تُنبَذ قيم السمة الحرجة وتُحابَى القيمة المتوسطة، فيثبت التجمع عليها، ممّا يقلل من التنوع الجيني ومن تفاوت القيم حول القيمة المتوسطة،[93][100] وذلك مثل أن يتقارب أفراد التجمع من نفس الطول المتوسط بمعدل بطيء.
الاصطفاء الجنسي هو حالة خاصة من الاصطفاء الطبيعي، وهو يحابي السمات التي تعزز من النجاح التناسلي (بالإنكليزية: reproductive success) للكائن الحي باستهوائها للجنس الآخر.[101] السمات التي تطورت بفعل الاصطفاء الجنسي تتجلى على الأخص في ذكور بعض فصائل الحيوانات. وذلك على الرغم من أن بعضها يقلل من فرص بقائهم، بما فيها القرون الثقيلة، نداءات التزاوج، والألوان الزاهية الملفتة للمفترسين.[102] إلا أنَّ نقطة الضعف هذه يُعَوض عنها بالنجاح التناسلي المُعَزَّز لدى الذكور الحاملين لتلك السمات المكلفة وصعبة التزييف، التي تم اصطفاؤها جنسياً.[103]
الانتقاء الطبيعي بشكل عام يجعل الطبيعة المعيار لتحديد أي من السمات تصلح أو لا تصلح للنجاة. تشير "الطبيعة " في هذا السياق إلى النظام البيئي، وهو النظام الذي يتفاعل فيه الكائن الحي مع العوامل الفيزيائية والأحيائية في بيئته المحلية. يوجين أودم (بالإنكليزية: Eugene Odum)، أحد مؤسسي علم البيئة، عرَّف النظام البيئي على أنه: "أي وحدة تحوي كل الكائنات الحية التي تعمل معاً في منطقة معينة تتفاعل مع البيئة الفيزيائية، بحيث يَنتُج عن تدفق الطاقة مبنى غذائي، تنوع حيوي، ودورات مادة (أي تبادل المواد بين العناصر الحيوية واللاحيوية) واضحي المعالم في النظام."[104] كل تجمع في النظام البيئي يحتل موقعاً بيئياً متمايزاً، ولديه علاقات متمايزة مع عناصر النظام الأخرى. هذه العلاقات تتضمَّن تاريخ حياة الكائن الحي، موقعه في السلسلة الغذائية، والنطاق الجغرافي الذي يمكن أن يعيش فيه. هذه الإحاطة الواسعة بالطبيعة تمكّن العلماء من تحديد القوى المعينة التي تشكل مجتمعة الانتقاء الطبيعي. يمكن أن يعمل الاصطفاء الطبيعي على مستويات تراتب مختلفة، مثل الجينات، الخلايا، أفراد الكائنات الحية، مجموعات الكائنات الحية والأنواع.[105][106][107] ويمكن أن يعمل الاصطفاء على عدة مستويات في آنٍ واحد.[108] إنَّ واحداً من الأمثلة على الاصطفاء الذي يحدث في مستوى دون الفرد الحي هو الينقولات، وهي الجينات الواثبة التي تستطيع التضاعف والانتشار في أنحاء الجينوم.[109] الاصطفاء الواقع على مستويات أعلى من الفرد، مثل الاصطفاء الزمري (بالإنكليزية: group selection)، قد يسمح بتطور تعاون بين الكائنات الحية، وقد تم تناول ذلك أدناه.
التطفر الانحيازي
بالإضافة لكونه مصدراً مهماً للتنوع الجيني، التطفّر (حدوث الطفرات) قد يعمل أيضاً كآلية تطور عندما تتفاوت احتمالات حدوث الطفرات المختلفة على المستوى الجزيئي. وهذا ما يُعرف بانحياز التطفّر. بعبارة أخرى: بعض الطفرات يكون حدوثها مرجح أو شائع أكثر من غيرها، ف"ينحاز" التطفر لها.[111] على سبيل المثال، إذا وجد نمطان جينيان يحملان نيوكليوتيدان لهما نفس الصلاحية ويقعان بمواقع متناظرة؛ الأول يحمل نيوكليوتيد G والآخر نيوكليوتيد A، لكن التطفّر(التبدّل) من G إلى A يحدث أكثر من التطفر من A إلى G، فعندها الأنماط الجينية ذات النيوكليوتيد A تميل أكثر لأن تتطور.[112] تباين الأصنوفات (بالإنكليزية: taxa) من حيث الانحياز لطفرات الغرز وطفرات الخبن، قد يؤدي لتطور أحجام مختلفة من الجينوم.[113][114] وقد لوحظ حدوث التطفر أو النماء الانحيازي في التطور على المستوى المورفولوجي أيضاً.[115][116] فمثلاً، طبقاً لنظرية التطور البالدويني، التطفر يمكن أن يؤدي بسمات كانت قد تطورت بتأثير البيئة لأن تتمثَّل جينيا.[117][118]
تأثيرات التطفر الانحيازي تتراكب على سيرورات أخرى، مثل الانتقاء. فإذا حابى الانتقاء طفرتين بقدر متساوٍ، لكن امتلاك كلتاهما معاً لا يعود بفائدة زائدة، فإنَّ الطفرة التي تحدث أكثر، على الأغلب هي التي ستترسخ في التجمع.[119][120] طفرات فقدان الوظيفة (التي تؤدي لفقدان وظيفة الجين) شائعة ومرجحة أكثر بكثير من الطفرات التي تنتج جينات جديدة وظيفية بالكامل.الانتقاء ينبذ معظم طفرات فقدان الوظيفة. ولكن إذا كانت قوة الانتقاء ضعيفة، فإنَّ انحياز التطفر لفقدان الوظيفة يمكنه التأثير على التطور.[121] مثلاً، الأخضاب (بالإنكليزية: pigments) تفقد فائدتها في حال انتقال الحيوانات للعيش في كهوف مظلمة، ومن ثم تميل لأن تُفقد.[122] هذا النوع من فقدان الوظائف يحدث بسبب انحياز التطفر، و/أو لأن لامتلاكها تكلفة، واختفاء فائدتها أدى بالانتقاء الطبيعي لأن ينبذها. لكن يبدو أن فقدان قدرة التبوغ لدى البكتيريا، خلال عمليات التطور المخبرية، سببه التطفر الانحيازي، لا الانتقاء الطبيعي ونبذه لتكلفة الاحتفاظ بهذه القدرة.[123] عندما لا يؤثر الانتقاء على فقدان الوظيفة، فإنَّ السرعة التي يتم بها الفقدان تكون متعلقة بمعدل التطفر (بالإنجليزية) أكثر منها بحجم التجمع الفعال (بالإنكليزية: effective population size)، [124] مما يشير إلى أنَّ فقدان الوظيفة مسيَّر بالتطفر الانحيازي أكثر من الانحراف الوراثي.
الانحراف الوراثي
الانحراف الوراثي (بالإنكليزية: genetic drift) هو تغير تواترات الألائل عشوائياً من جيل لآخر بسبب التوزيع العشوائي للألائل. أي أنَّ الألائل تخضغ لخطأ الاستعيان (بالإنكليزية: sampling error)، وهو الخطأ في تمثيل العينة للمجتمع. فالألائل التي يحصل عليها النسل (العينة)، تكون تواتراتها مختلفة عن تواترات ألائل الجيل السلفي (المجتمع).[125] فعندما تكون القوى الانتقائية ضعيفة نسبياً أو معدومة، "ينحرف" تواتر الألائل لأعلى وأسفل على نحو عشوائي (بسير عشوائي). ويتوقف التواتر عن الانحراف حين يثبت الأليل - إما يزول من التجمع أو يستبدل الألائل الأخرى بالكامل. لهذا قد يتسبب الانحراف الوراثي بإزالة ألائل من التجمع نتيجة للصدفة فقط. حتى بغياب القوى الانتقائية، الانحراف الوراثي قد يؤدي لانحراف وتباعد تجمعين منفصلين كانا قد بدآ بنفس المبنى الجيني، إلى أن يصبحا تجمعين مختلفين بمجموعات مختلفة من الألائل.[126]
عادةً يصعب قياس مدى أهمية الأدوار التي يلعبها الاصطفاء والسيرورات المحايدة، مثل الانحراف الجيني، كقوى محركة للتطور.[127] إن الأهمية النسبية لكل من القوى التكيفية وغير التكيفية في تحريك التغير التطوري هي إحدى المواضيع التي لا تزال قيد البحث في علم الأحياء التطوري.[128]
وفقاً لنظرية التطور الجزيئي المحايدة، معظم التغيّرات التطورية هي ناجمة عن تثبيت الطفرات المحادية بفعل الانحراف الجيني.[5] وبحسب هذا النموذج، هذا يعني أنَّ معظم التغيّرات الوراثية في التجمع هي ناتجة عن ضغط التطفّر المستمر والانحراف الجيني.[129] هذه الصيغة من النظرية المحايدة أصبحت في الغالب مهجورة الآن لأنها لا تتوافق مع التنوع الجيني الملاحظ في الطبيعة.[130][131] إلا أنَّ هناك صيغة حديثة ومدعومة أكثر لهذا النموذج، ألا وهي النظرية شبه المحايدة، وبموجبها الطفرة التي تكون محايدة في تجمع صغير، قد لا تكون كذلك في تجمع آخر أكبر.[93] وهناك نظريات بديلة أخرى تشير إلى أنَّ الانحراف الجيني تضعفه قوى تطورية عشوائية أخرى، مثل الترافق الجيني، الذي يعرف أيضاً بالجرّ الجيني.[125][132][133] الزمن اللازم لثبات أليل محايد بواسطة الانحراف الجيني يتناسب طردياً مع حجم التجمع.[134] إلا أنَّ عدد أفراد التجمع ليس مهماً هنا، وإنما حجم التجمع الفعال هو المقياس الأساسي.[135] التجمع الفعال عادةً يكون أصغر حجماً من التجمع الكلي بما أنَّه بالأخير هناك عوامل تؤخذ بالحسبان، مثل درجة التوالد الداخلي (زواج الأقارب)، ومرحلة دورة الحياة التي يكون حجم التجمع فيها أصغر ما يمكن.[135] وحجم التجمع الفعال قد يختلف بالنسبة لكل جين في نفس التجمع.
الترافق الجيني
التأشيب (بالإنكليزية: recombination) يسمح بتفرُّق الألائل المتواجدة على نفس شريط الدنا. لكن بما أن معدل التأشيب منخفض (تقريباً يحدث مرتين في كل كروموسوم في كل جيل)، فإنَّ الجينات التي تتموضع في مواقع متقاربة على الكروموسوم ينخفض احتمال تفرقتها، ويرتفع احتمال توريثها سوياً، وهذه الظاهرة تعرف بالارتباط الجيني (بالإنكليزية: linkage). [137] إحتمال توريث أليلان (شكلان مختلفان لنفس الجين) متقاربان سوياً يُحَدَّد عبر قياس اختلال توازن ارتباطهم (بالإنكليزية: linkage disequilibrium)، وهو الفرق بين عدد مرات ظهورهما معاً في الكروموسوم وبين عدد المرات المتوقع بناءً على مقدار تواتر كل منهما. مجموعة الألائل المتجاورة التي عادةً ما تُوَرَّث سوياً تُسَمَّى بالنمط الفرداني (بالإنكليزية: haplotype). هذا الأمر مهم عندما يكون أحد الألائل من بين المجموعة نافعاً جداً، إذ أنَّ الاصطفاء الطبيعي، بمحاباته لهذا الأليل، قد يسبب مسحاً اصطفائياً (بالإنكليزية: selective sweep)، فيه تصبح باقي ألائل النمط الفرداني شائعة في التجمع، وهذه الظاهرة تُعْرَف بالترافق الجيني أو الجر الجيني (بالإنكليزية: genetic hitchhiking). [138] وحجم ملائم من التجمع الفعال يتأثَّر جزئياً بالجر الجيني الذي يحدث من جراء ارتباط جينات محايدة بجينات أخرى وقع عليها الاصطفاء الطبيعي.
انسياب الجينات
انسياب الجينات هو تبادل الجينات بين التجمعات أو الأنواع المختلفة.[85] وجود أو غياب انسياب الجينات يغيّر من مسار التطور بشكل جذري. وبسبب تعقيد الكائنات الحية، فإنَّ التجمعين المعزولين تماماً عن بعضهما سيتطوران بشكل مختلف ومتفاوت تماما عن بعضهما، حتى وإن بقي التجمعان متشابهان من حيث تكيفهما مع المحيط.
إذا ظهر اختلاف جيني بين التجمعات، فإنَّ انسياب المورثات بينها قد يُدخل سمات أو ألائل ضارة بالنسبة لكل من التجمعات (بما أنها دخيلة وليست متوافقة مع بيئتهم)، الأمر الذي قد يؤدي بالكائنات الحية ضمن كل تجمع لأن تُطوِّر آليات تمنع تزاوجها مع التجمعات البعيدة عنها جينياً، مما يتسبب مع الوقت في ظهور أنواع جديدة. ولذلك يُعدّ تبادل المعلومات الوراثية بين الأفراد أمراً مهماً جداً لتطور مفهوم النوع الأحيائي (BSC). أثناء تطوير الاصطناع التطوري الحديث، طوَّر سيوال رايت نظرية توازن التناوب (بالإنجليزية) الخاصة به، التي تقول بأنَّ انسياب المورثات بين التجمعات المعزولة جزئياً كان عاملاً مهماً في التطور التكيفي.[139] بيد أن أهمية هذه النظرية أصبحت تلقى انتقادات شديدة مؤخراً
النتائج
التطور يؤثر على كل جانب من سلوكيات الكائنات الحية ومظاهرها. ومن أبرزها التكيفات السلوكية والفيزيائية الناتجة عن عملية الاصطفاء الطبيعي. هذه التكيفات تزيد من صلاحية الكائن الحي عبر تحسين قدرة العثور على الطعام، تجنب الحيوانات المفترسة، أو اجتذاب الأزواج. تتفاعل الكائنات الحية مع عملية الاصطفاء أحياناً بتشكيل علاقات تعاونية مع بعضها البعض، وذلك عادةً عن طريق مساعدة الأقارب أوالتعايش النفعي المتبادل. على المدى الطويل، يُنتج التطور أنواع جديدة عبر فصل التجمعات السلفية عن بعضها البعض، لتكون تجمعات جديدة تنتهي لأن تصبح غير قادرة على التزاوج فيما بينها.
نتائج التطور هذه تُصنف أحياناً لفئتين: تطور كبروي (بالإنكليزية: macroevolution) وتطور صغري (بالإنكليزية: microevolution). هذا الأول هو تطور واسع النطاق يحدث في أو فوق مستوى النوع، كالانقراض والانتواع. والأخير هو تغيرات تطورية أصغر تحدث ضمن النوع (بين أفراد النوع أو بين تجمعاته)، مثل التكيفات.[141] عموماً يُعتبر التطور الكبروي نتيجة لتراكم تأثيرات التطور الصغري على مدى فترات طويلة.[142] إذاً، الفرق بين التطور الكبروي والصغري ليس جوهرياً - فهو ببساطة يكمن بالمدة الزمنية المستغرقة.[143] غير أنَّ في التطور الكبروي، سمات النوع بأكمله قد تكون مهمة، بما أنها قد تؤثر على احتمال حدوث الانتواع أو الانقراض. فمثلا وجود تنوع كبير في النوع يسمح بتكيفه بسرعة مع المواطن الجديدة، مما يقلل من احتمال انقراضه، بينما النطاق الجغرافي الواسع يزيد من احتمال الانتواع، وذلك بزيادة احتمال انعزال جزء من التجمع عن التجمع الأصلي. هنا، قد يدخل في التطور الكبروي والصغري عمل الاصطفاء الطبيعي على مستويات مختلفة - بحيث يعمل في التطور الصغري على مستوى الجينات والكائنات الحية، فيما عمليات التطور الكبروي، مثل اصطفاء الأنواع، تعمل على أنواع بأكملها وتؤثر على معدلات انتواعها وإنقراضها.[144][145][146] هناك إعتقاد شائع خاطئ بأن للتطور أهداف أو خطط بعيدة المدى، ولكنه في واقع الأمر ليس له أي أهداف على المدى البعيد، وليس من الضرورة أن يّنتج عنه زيادة في التعقيد.[147][148] مع أن الكائنات الأكثر تعقيداً قد نتجت عن عملية التطور، إلا أنَّ ذلك مجرد ناتج عرضي لازدياد العدد الإجمالي للكائنات الحية، وأشكال الحياة الأقل تعقيدا ما زالت هي الشائعة أكثر في الغلاف الحيوي.[149] فمثلاً الغالبية الساحقة من الأنواع تؤلفها بدائيات النواة المجهرية، والتي تشكل، رغم حجمها الصغير، حوالي نصف الكتلة الحيوية،[150] وهي تشكل أيضاً الغالبية العظمى من التنوع الحيوي على كوكب الأرض.[151] لذا فإنَّ أشكال الحياة البسيطة على مر التاريخ كانت وما زالت هي صورة الحياة السائدة على كوكب الأرض، وأشكال الحياة المعقدة تبدو أكثر شيوعاً فقط لأنها بارزة أكثر.[152] إنَّ تطور الكائنات الحية الدقيقة مهم جداً للبحوث التطورية الحديثة، حيث أن تكاثرها السريع يسمح بدراسة تطورها مخبرياً (بالإنجليزية) وبملاحظة التطور والتكيف خلال الوقت الفعلي.
التكيف
التكيف هو العملية التي تصبح فيها الكائنات الحية ملائمة أكثر للعيش في موطنها.[155][156] مصطلح "التكيف" قد يُستخدم أيضاً للإشارة لسمة فيزيائية أو سلوكية نتجت عن عملية التكيف لتحسن من قدرة الكائن الحي على المحافظة على حياته وبقاء جنسه. مثال على ذلك هو تكيف أسنان الحصان على طحن العشب - وقدرته على طحن العشب بأسنانه المترافعة هي تكيف. للتمييز بين هذين الاستخدامين للمصطلح، يُستعمل مصطلح "التكيف" للإشارة للعملية التطورية، والمصطلح "صفة تكيفية" للدلالة على السمات الناتجة عن العملية التطورية. الصفات التكيفية هي ناجمة عن الاصطفاء الطبيعي.[157] وقد عيَّن عالم الأحياء التطورية ثيودوسيوس دوبجانسكي التعريفات التالية:
- التكيف هو العملية التطورية التي تتحسن بها قدرة الكائن الحي على العيش في موطنه أو مواطنه.[158]
- التكيفية هي حالة التكيف التي يكون عليها الكائن الحي، أي الدرجة التي يستطيع فيها الكائن الحي العيش والتكاثر في مواطن معينة.[159]
- الصفة التكيفية هي جانب من نمط الكائن الحي النمائي، والتي تُوْجِد أو تحسن من فرص بقائه وتكاثره.[160]
التكيف قد يتسبب إما باكتساب خصائص جديدة أو بفقدان خصائص سلفية. من الأمثلة التي يظهر فيها كلا النوعين من التغييرات هو تكيف البكتيريا مع الضغط الاصطفائي الذي تفرضه المضادات الحيوية عليها، إذ تحصل بالبكتيريا تغييرات جينية تكسبها مقاومة لها، وذلك عن طريق إحداث تغيير بمستهدفات الدواء، و/أو عبر زيادة نشاط الناقلات التي تضخ الدواء لخارج الخلية.[161] بعض الأمثلة البارزة الأخرى هي: تطوير الإشريكيات القولونية E. coli للقدرة على استخدام حمض الستريك كمصدر غذاء في تجربة مخبرية طويلة الأمد،[162] وأيضاً تطوير الصيفرية (بالإنكليزية: Flavobacterium) لإنزيم جديد كلياً يسمح لها بالتغذي على المخلفات الناتجة عن تصنيع النايلون،[163][164] وبالإضافة لبكتيرات التربة السفينغوبيوم (بالإنكليزية: Sphingobium) والتي طورت مساراً استقلابياً جديداً لتحليل مبيد الآفات الاصطناعي بنتاكلوروفينول (بالإنكليزية: pentachlorophenol). [165][166] من الأفكار المثيرة للاهتمام والجدل هي أن بعض التكيفات قد تزيد من قدرة الكائنات الحية على توليد التنوع الجيني والتكيف بواسطة الاصطفاء الطبيعي (أي أنها تزيد من تطورية الكائن الحي).
يحدث التكيف عن طريق حدوث تعديلات تدريجية في البنى الموجودة. لذلك البنى المتشابهة في تنظيمها الداخلي قد تكون لها وظائف مختلفة في الفصائل المتقاربة. فهذه هي نتيجة تكييف بنية سلفية واحدة على العمل بطرق مختلفة. على سبيل المثال، عظام أجنحة الخفافيش تشبه كثيراً تلك التي في أقدام الفئران وأيدي الرئيسيات، وذلك يعود لانحدار كل هذه البنى من سلف ثديي مشترك.[172] ولكن بما أنَّ كل الكائنات الحية تربط بينها قرابة إلى حد ما،[173] فإنَّه حتى الأعضاء التي يبدو الشبه بينها قليلاً أو معدوماً، مثل أعين كل من المفصليات والرخويات والفقاريات، أو أطراف وأجنحة المفصليات والفقاريات، قد تكون مستندة إلى مجموعة مشتركة من الجينات المتماثلة، التي تتحكم بتراكيبها ووظائفها. وهذا ما يُسمَّى بالتماثل العميق[174][175]
خلال التطور، قد تفقد بعض البنى وظائفها الأساسية وتصبح بالتالي بنى آثارية.[176] وظائف هذه البنى قد تكون ضئيلة أو معدومة في النوع الحالي، ولكن تكون لديها وظائف واضحة المعالم في نوع سلفي أو في نوع آخر على درجة كبيرة من القرابة. تتضمَّن الأمثلة على ذلك: الجينات الكاذبة،[177] آثار الأعين اللاوظيفية في الأسماك العمياء التي تعيش في الكهوف،[178] أجنحة الطيور التي لا تطير،[179] وتواجد عظام حوض في الأفاعي والحيتان.[171] الأمثلة على البنى اللاوظيفية في الانسان (بالإنجليزية) تتضمن: أضراس العقل،[180] العصعص،[176] الزائدة الدودية،[176] وبعض السلوكيات اللاوظيفية مثل تقبب الجلد (بالإنكليزية: goose bumps)،[181][182] وردود الأفعال البدائية (بالإنجليزية). [183][184][185]
غير أنَّ الكثير من السمات التي تبدو وكأنها تكيفات بسيطة هي في الحقيقة تكانفات، أي تكيفات آنفة (سابقة) (بالإنكليزية: exaptations). وهي بنى كانت قد تكيفت لتأدية وظيفة معينة، ولكنها بالمصادفة أصبحت مفيدة لوظيفة أخرى خلال التطور.[186] إنَّ واحداً من الأمثلة على ذلك هو السحلية الأفريقية المنزلقة (بالإنكليزية: Holaspis guentheri)، التي كانت قد طورت رأساً شديد التفلطح يمكّنها من الاختباء في الصدوع، كما يُلاحظ عند أقربائها الأقربين. ولكن في هذا النوع تحديداً، أصبح الرأس مسطحا لدرجة أنه يساعدها على الانزلاق الهوائي من شجرة لشجرة،[186] لذلك يُعتبر رأسها المسطح تكانفاً. ومن الأمثلة الأخرى هي بعض الآلات الجزيئية المتواجدة في الخلايا، والتي تطورت عبر توظيف عدة بروتينات موجودة سابقا كانت تقوم بوظائف أخرى مختلفة،[141] مثل الأسواط البكتيريّة،[187] وآلات فرز البروتينات.[188]
مثال إضافي آخر هو توظيف انزيمات حالَّة للسكر وأخرى أيضية لتعمل كبروتينات بنائية تُسَمَّى غلوبولينات العدسة (بالإنجليزية)، وهي موجودة في العدسة العينية للمتعضيات.[189][190]
أحد المبادئ الحساسة في علم البيئة هو لاقصاء التنافسي، وينص على أنه لا يمكن لأي نوعين أن يحتلا نفس الموضع في نفس البيئة لفترة زمنية طويلة.[191] وبالتالي يميل الاصطفاء الطبيعي لإجبار الأنواع على التكيف للعيش في مواضع بيئية مختلفة. هذا قد يعني مثلاً، أنَّ نوعين معينين من سمك السكليد قد يتكيفا للعيش بمواطن مختلفة، مما سيقلل من التنافس بينهما من أجل الغذاء، ويجنبهما الإقصاء.[192]
من المجالات التي تخضع للدراسات حاليا في علم الأحياء التطوري النمائي هي الأسس التنموية التي تقوم عليها التكيفات والتكانفات (تكيفات آنفة).[193] هذا البحث يتناول منشأ وتطور عملية النمو الجنيني، وكيف تنشأ عن التعديلات على النمو والعمليات التنموية مميزات وخصائص جديدة.[194] وأظهرت هذه الدراسات أنَّ التطور قد يعدل من النمو، مما يجعله ينتج بنى جديدة. على سبيل المثال، بنى العظام الجنينية التي تتطور وتشكل الفك في الفقاريات غير الثديية، تتطور لتشكل بدلاً من ذلك جزءا من الأذن الوسطى في الثديات.[195] بعض البنى التي فقدت خلال التطور، من الممكن أن تعود لتظهر ثانيةً بسبب تغييرات تطرأ على الجينات التنموية، كالطفرة في الدجاج التي تجعل الأجنة تنمي أسناناً تشبه تلك لدى التماسيح[196] وغدا يتضح الآن أنَّ معظم التعديلات التي تطرأ على أشكال الكائنات الحية تعود إلى تغييرات تحدث في مجموعة صغيرة من الجينات المحافظة.
التطور المشترك
التآثرات (بالإنكليزية: interactions) بين الكائنات الحية ينتج عنها صراع وأيضاً تعاون. وعندما يكون التفاعل بين أزواج من الأنواع، كما في حال الممراض والمضيف، أو المفترس والفريسة، فإنَّها قد تطور مجموعات متلائمة من التكيفات. بحيث يؤدي تطور نوع ما لحدوث تكيفات في نوع ثان، والتي تؤدي بدورها لحدوث تكيفات في النوع الأول. هذه الحلقة من الانتخاب الطبيعي والاستجابة له تُسَمَّى بالتطور المشترك (بالإنكليزية: co-evolution). [198] مثال على ذلك هو إنتاج سم الينفوخ في سمندل الماء خشن الجلد (بالإنجليزية)، وتطوير مقاومة لهذا السم في مفترسه، الأفعى المقلمة (بالإنجليزية). وأدى سباق التسلح التطوري في هذا الزوج المؤلف من فريسة ومفترس لإنتاج مستويات عالية من السم في السمندل، وبالمقابل صاحَبَه تطور مستويات عالية من مقاومة للسم في الأفعى.
التعاون
ليست كل التآثرات التي تتطور بشكل مشترك بين الأنواع تنطوي على صراع.[200] فقد تطورت الكثير من التآثرات النفعية المتبادلة. على سبيل المثال، ثمة تعاون شديد بين النباتات والفطريات الجذرية، التي تنمو على جذورها وتساعدها على امتصاص المغذيات من التربة.[201] وهذه العلاقة تبادلية (بالإنجليزية)، إذ تقوم النباتات بتزويد الفطريات بالسكريات الناتجة عن التمثيل الضوئي. إنَّ الفطريات هنا تنمو داخل خلايا النباتات، مما يتيح لها تبادل المغذيات مع مضيفها، وبنفس الوقت إرسال إشارات تثبط جهاز النبات المناعي.[202]
وقد تطورت تحالفات بين أفراد ينتمون لنفس النوع أيضا. ومن الحالات القصوى هي الاجتماعية العليا (بالإنكليزية: eusociality) - سلوك إجتماعي على درجة عالية من التنظيم - الموجودة عند الحشرات الاجتماعية مثل النحل، النمل، والأرضة، حيث تقوم الحشرات العقيمة بحماية وإطعام القلة القادرة على التكاثر في المستعمرة. ومن الأمثلة على التحالفات التي تحدث على نطاق أصغر، هي الخلايا الجسدية المُكونة لأجساد الكائنات الحية، إذ تحافظ ضوابط تكاثرها على على استقرار الكائن الحي، الذي بدوره ينتج خلايا جنسية لانجاب ذرية. هنا تستجيب الخلايا الجسدية لإشارات محددة تأمرها إما بأن تنمو، تبقى على حالتها، أو تموت. وإن لم تستجب لهذه الأوامر وتضاعفت بدون ضوابط، فإن نموها المفرط سيتسبب بالسرطان.[203] من المحتمل أنَّ مثل هذا التعاون بين أفراد نفس النوع قد تطور بواسطة اصطفاء القرابة (بالإنكليزية: kin selection)، الذي يقوم فيه أحد أفراد النوع بمساعدة أقربائه على التكاثر.[204] يحابي الانتقاء المساعدة بين الأقارب، وذلك لأنه إن تواجدت ألائل معززة لسلوك المساعدة في الفرد المُساعد، فمن المرجح أن يكون أقرباءه حاملين لها أيضا، ولذلك تجد هذه الألائل طريقها إلى الجيل التالي.[205] إحدى العمليات الأخرى المعززة للتعاون هي الاصطفاء الزمري، فيه تعود فائدة التعاون على مجموعة من الكائنات الحية.
الانتواع
الانتواع هو العملية التي يتشعب فيها النوع إلى نوعين سليلين أو أكثر.[207]
ثمة طرق مختلفة لتعريف مفهوم النوع. واختيار التعريف يعتمد على خصائص النوع المعنى بها.[208] فمثلاً بعضها ينطبق أكثر على الكائنات الحية التي تتكاثر جنسياً، فيما البعض الآخر يكون لاستخدامه فائدة أكثر عند التعامل مع الكائنات التي ليس لديها تكاثر جنسي. ورغم تنوع التعريفات الكبير، إلا أنه يمكن تصنيف كل منها ضمن أحد التوجهات الفلسفية الثلاثة الشاملة: البيئي، التزاوجي البيني، والتطور السلالي.[209] مفهوم النوع الأحيائي هو مثال مثال كلاسيكي على توجه التزاوج البيني. ففي عام 1942، قدَّم إرنست ماير مفهوم النوع الأحيائي الذي عرَّف النوع على أنه: مجموعة من التجمعات التي لها القدرة على التزاوج فيما بينها، والمعزولة تزاوجياً عن المجموعات الأخرى.[210] وعلى الرغم من استعمال مفهوم النوع الأحيائي الواسع وطويل الأمد، إلا أنَّ به ما يثير الجدل كغيره من التوجهات التزاوجية البينية، وأحد الأسباب هو أنها لا يمكن تطبيقها على بدائيات النوى، التي تفتقد للتكاثر الجنسي.[211] وهذا ما يعرف بمشكلة النوع.[208] وقد حاول بعض الباحثين الوصول لمفهوم واحد موحد، بينما تبنى آخرون توجها تعددياً واقترحوا انه قد تكون هناك عدة طرق مختلفة لتفسير تعريف النوع بشكل منطقي.[208][209]
وجود حواجز تكاثرية بين تجمعين جنسيين متفرعين من نوع واحد هو شرط ضروري كي يصبحا نوعان جديدان. الانسياب الجيني قد يبطيء من هذه العملية بنقله للاختلافات الجينية الجديدة إلى التجمعات الأخرى. تمكُّن الأنواع من التزاوج فيما بينها يعتمد على مدى التباعد الذي حدث بينها منذ تحدرها من سلفها المشترك الأخير، فهناك أنواع تستطيع انجاب نسل، مثل الأحصنة والحمير، التي ينتج عن تزاوجها البغل.[212] عادةً يكون النسل الهجين الناتج عقيما. فرغم أن بعض الأنواع ذات القرابة الكبيرة قد تكون قادرة على التزاوج بانتظام، إلا أن الاصطفاء ينبذ نسلها الهجين، وتبقى هذه الأنواع متمايزة. غير أنه أحيانا قد تنتج هجائن قادرة على التكاثر والبقاء، وهذه الأنواع الهجينة الجديدة قد تمتلك إما خصائص وسطية ناتجة عن مزج خصائص نوعي الأبوين، أو قد تكون لها خصائص ظاهرية جديدة.[213] إنًّ مدى أهمية التهجين في إنشاء أنواع جديدة ما زال غير واضح. إلا أنه شوهدت حالات تهجين في الكثير من الحيوانات، [214] بما فيها ضفدع الشجر الرمادي، الذي يعتبر حالة مدروسة جيدا.[215]
لقد دُرس الانتواع مرات عديدة في ظروف مخبرية مضبطة وكذلك في الطبيعة.[216] الانتواع الذي يحدث في التجمعات التي تتكاثر جنسيا يكون سببه العزل التكاثري المتبوع بالتباعد السلالي. هنالك أربع آليات يحدث بها الانتواع، وأكثرها شيوعاً لدى الحيوانات هو الانتواع التبايني (بالإنكليزية: allopatric speciation). وهو يحدث في تجمعات عُزلت جغرافياً، الأمر الذي قد يَحدث بفعل الهجرة أو العمليات الجيولوجية المسببة لتجزؤ الموطن. وفي هذه الظروف، يُحدث الاصطفاء الطبيعي تغيرات سريعة في مظاهر الكائنات الحية وسلوكياتها.[217][218] وبما أنَّ التجمع المنعزل عن التجمع الأصلي يتعرض للانتقاء الطبيعي والانجراف الجيني بشكل مستقل ومختلف عن بقية تجمعات النوع، فإنَّ الانعزال الجغرافي قد ينتج عنه بنهاية المطاف كائنات غير قادرة على التزاوج فيما بينها.[219]
آلية الانتواع الثانية هي الانتواع الخارجي (بالإنكليزية: peripatric speciation)، وهو يحدث عندما تُفصل تجمعات صغيرة عن التجمع الأصلي في بيئة خارجية جديدة. وهو يشبه الانتواع التباعدي من حيث الانعزال الجغرافي والتكاثري، لكن بخلافه، في الانتواع الخارجي يكون حجم التجمع المفصول أصغر بكثير من حجم التجمع الأصلي. وهنا يؤدي تأثير المؤسس لحدوث انتواع سريع، بعد أن يؤدي تزايد التوالد الداخلي لزيادة الضغط الانتقائي على تماثل الزيجوت، الأمر الذي يؤدي إلى تغير جيني سريع.[220]
آلية الانتواع الثالثة هي الانتواع المحاذي (بالإنكليزية: parapatric speciation)، وهو يشبه الانتواع الخارجي من حيث انعزال مجموعة صغيرة في موطن جديد، إلا أنه لا يوجد هنا حاجز فيزيائي يعزل بين التجمعين. وإنما ينتج الانتواع عن تطور آليات تقلل من انسياب الجينات فيما بين التجمعين.[207] 205 بشكل عام، هذا يحدث نتيجة لحصول تغييرات بيئية جذرية في موطن النوع. مثال على ذلك هو العشب الربيعي (بالإنكليزية: Anthoxanthum odoratum) الذي يمر بالانتواع المحاذي استجابةً للتلوث المعدني الموضعي الصادر عن المناجم.[221] فهنا تطورت نباتات مقاومة لنسبة المعادن العالية في التربة. والاصطفاء بنبذه للتزاوج مع التجمع الأصلي الحساس للمعادن أدى لإحداث تغيير تدريجي في وقت إزهار النباتات المقاومة للمعادن، الأمر الذي أفضى إلى نشوء عزل تكاثري كامل. الاصطفاء بنبذه للتهجين بين التجمعين قد يؤدي إلى التدعيم، أي تطور سمات تعزز من التزاوج ضمن نفس النوع، وأيضاً لانزياح الخواص، أي زيادة التمايز الظاهري بين النوعين.[222]
والآلية الرابعة هي الانتواع التماثلي (بالإنكليزية: sympatric speciation). في هذا الانتواع يتشعب النوع دون حدوث عزل جغرافي أو تغيير في الموطن. هذه الآلية نادرة، وذلك لأن أدنى انسياب للمورثات قد يلغي الاختلاف الجيني في التجمع.[223] بشكل عام، يتطلب حدوث هذا الانتواع في الحيوانات تطور اختلافات جينية بالإضافة لتزاوج غير عشوائي من أجل السماح للعزل التكاثري بالتطور.[224]
أحد أنواع الانتواع التماثلي يتضمن حدوث تزاوج ما بين نوعين قريبين لإنتاج نوع هجين جديد. هذا ليس شائع الحدوث، إذ أن الحيوانات المهجنة بالعادة تكون عقيمة. وهذا يرجع إلى أنَّه خلال الانتصاف، تكون الكروموسومات المثلية لكل من الأبوين تابعة لنوعين مختلفين، ولا يمكنها الازدواج. ولكن هذا الأمر شائع أكثر في النباتات، وذلك لأن النباتات عادة تُضاعف عدد كروموسوماتها، مما يجعلها متعددة الصيغ الكروموسومية. [225] فهذا يسمح للكرموسومات الصادرة عن كل من النوعين الأبويين بتشكيل أزواج متلائمة خلال الانتصاف، بما أن كروموسومات الأبوين هي بالأصل مُمَثلة بأزواج.[226] إنَّ أحد الأمثلة على هذا النوع من الانتواع هو تزواج النوعين أرابيدوپسس ثاليانا (بالإنكليزية: Arabidopsis thaliana) وأرابيدوپسس أرينوزا (بالإنكليزية: Arabidopsis arenosa) لينتجا النوع الجديد أرابيدوپسس سويسيكا (بالإنكليزية: Arabidopsis suecica). [227] وهذا قد حدث قبل حوالي 20000 سنة،[228] وأعيدت عملية الانتواع هذه مخبرياً، مما أتاح دراسة الآلية الوراثية المتعلقة بهذه العملية.[229] إنَّ تضاعف الكروموسومات قد يكون مسببا شائعا للانعزال التكاثري، إذ أنَّ نصف الكروموسومات المضاعفة لن تزدوج أثناء التكاثر مع الكائنات ذات الكروموسومات غير المضاعفة.[230] أحداث الانتواع مهمة في نظرية التوازن المتقطع (بالإنجليزية)، والتي تفسر النمط في السجل الأحفوري للهبَّات القصيرة من التطور والموزعة على فترات طويلة من الركود، حيث تبقى الأنواع ثابتة نسبياً دون تغيير.[231] في هذه النظرية، يكون الانتواع والتطور السريع مترابطين، فيما يعمل كل من الانتقاء الطبيعي والانحراف الجيني على الكائنات الحية التي تمر بالانتواع في مواطن جديدة أو تجمعات صغيرة. ولذلك فترات الركود في السجل الأحفوري تلائم التجمع الأصلي، أما الكائنات التي تمر بانتواع وتطور سريع تكون موجودة في تجمعات صغيرة ومقيدة جغرافياً، ولذلك تأحفرها يكون نادراً.
الانقراض
الانقراض هو اختفاء نوع بأكمله. الانقراض ليس حدثا غير اعتيادي، فالأنواع بشكل مستمر تظهر بالانتواع وتختفي بالانقراض.[233] تقريبا جميع أنواع الحيوانات والنباتات التي عاشت على سطح الأرض هي الآن منقرضة،[234] ويبدو أن الانقراض هو المصير النهائي لكافة الأنواع.[235] هذه الانقراضات حدثت كثيرا عبر تاريخ الحياة، إلا أن معدل الانقراض قد يرتفع في الانقراضات الجماعية الأحيائية.[236] انقراض العصر الطباشيري-الثلاثي، الذي انقرضت فيه الديناصورات غير الطائرة، هو الانقراض المعروف أكثر. ولكن انقراض العصر البرمي الترياسي كان أكثر شدة، إذ انقرض فيه تقريبا 96% من الأنواع.[236] انقراض العصر الهولوسيني (بالإنجليزية) هو انقراض جماعي جارٍ مرتبط بتوسع البشرية عبر الكرة الأرضية خلال عدة آلاف السنوات الماضية. معدلات الانقراضات الحالية هي أعلى ب-100-1000 مرة من قبل، وحوالي 30% من الأنواع الحالية قد تصبح منقرضة مع حلول منتصف القرن الواحد والعشرين.[237] نشاطات الإنسان هي الآن المسبب الرئيسي للانقراض المستمر،[238] وقد يعجل الاحترار العالمي من العملية في المستقبل.[239] دور الانقراض في التطور ليس مفهوماً بشكل جيد، وقد يتعلق الأمر بنوع الانقراض المعنى به.[236] مسببات الانقراضات الصغيرة المتواصلة، والتي تمثل معظم الانقراضات، قد تكون ناتجة عن تنافس الأنواع من أجل الموارد المحدودة (الاقصاء التنافسي).[47] فإن تمكَّن نوع ما من التفوق على نوع آخر، قد ينتج عن ذلك اصطفاء على مستوى الأنواع، حيث ينجو النوع الأصلح وتقاد باقي الأنواع إلى الفناء.[106] الانقراضات الجماعية المتقطعة هي أيضا مهمة، ولكن بدل أن تعمل كقوة اصطفائية، فإنها تقلل من التنوع بشكل كبير وبطريقة عشوائية، وتشجع حدوث هبَّات من التطور السريع وانتواعات في الكائنات الناجية.
تاريخ الحياة التطوري
أصل الحياة :
يُعتقد أنَّ كيمياء طاقية جداً أدت لإنتاج جزيء ذاتي التناسخ قبل حوال 4 بلايين سنة، وأن بعد نحو نصف بليون سنة عاش آخر سلف مشترك للحياة كلها.[241] وهناك إتفاق علمي على أنَّ الكيمياء الحيوية المعقدة المشكلة للحياة نتجت عن تفاعلات كيميائية أبسط.[242] من الممكن أنَّه كانت في بدء الحياة جزيئات ذاتية الاستنساخ مثل الرنا،[243] بالإضافة إلى جمعية الخلايا البسيطة.
السلف المشترك
الكائنات الحية على الأرض كلها متحدرة من سلف مشترك أو تجميعة جينات سلفية.[173][245] الأنواع الموجودة حالياً هي من مراحل عملية التطور، وتنوعها هو نتيجة لسلسلة طويلة من الانتواعات والانقراضات.[246] تم الاستدلال على السلف المشترك من أربع حقائق بسيطة تخص الكائنات الحية: أولاً، للكائنات الحية توزيعات جغراية لا يمكن عزوها للتكيف المحلي. ثانياً، تنوع الحياة ليس بتشكيلة من الكائنات الفريدة كلياً، وإنما من كائنات بينها تشابهات مورفولوجية. ثالثاً، السمات الأثارية التي تفتقد لوظائف واضحة تشابه سمات سلفية وظيفية. وأخيراً، يمكن تصنيف الكائنات الحية في تراتب من المجموعات المتداخلة بالاستناد إلى هذه التشابهات - على غرار شجرة العائلة.[247] ولكن اقترح بحث حديث أنَّه بسبب نقل الجينات الأفقي، قد تكون "شجرة الحياة" هذه أكثر تعقيداً من مجرد شجرة متفرعة بسيطة بما أنَّ هناك جينات تنتقل بمعزل عن التوريث بين أنواع بعيدة القرابة.[248][249]
أيضاً الأنواع السالفة تركت سجلات لتاريخها التطوري. فالأحافير تشكل مع التشريح المُقارَن للكائنات الحالية السجل المورفولوجي أو التشريحي.[250] وعن طريق مقارنة البنى التشريحية للأنواع الحديثة والمنقرضة، يستطيع البليونتولوجيين الاستدلال على أصول هذه الأنواع. إلا أنَّ هذه الطريقة تكون أكثر نجاحاً عند التعامل مع كائنات حية ذات أعضاء جسدية صلبة مثل الصدف، العظام والأسنان. فضلاً عن أنَّ بدائيات النواة مثل البكتيريا والعتائق لديها قدر محدود من المورفولوجيات المشتركة، ولذلك أحافيرها لا تزود معلومات عن أصلها. وفي الآونة الأخيرة أتت أدلة على السلف المشترك من دراسة التشابهات الكيميائية الحيوية بين الكائنات الحية. فمثلاً كل الخلايا الحية تستعمل نفس المجموعة الأساسية من النوكليوتيدات والأحماض الأمينية.[251] وكشف علم الوراثة الجزيئي مع تقدمه وجود سجل للتطور باقٍ في الجينوم الخاص بالكائن الحي: قياس التاريخ الذي تفرعت به الأنواع بواسطة تقنية الساعات الجزيئية التي تنتجها الطفرات.[252] على سبيل المثال، أسفرت هذه المقارنات بين تسلسلات الدنا عن أنَّ الإنسان والشمبانزي يتطابقان في 96% من الجينوم، وتحليل مناطق الاختلاف يساعد على تبيان التاريخ الذي عاش به السلف المشترك الذي انشق عنه هذان النوعان.
تطور الحياة
عاشت بدائيات النوى على الأرض منذ نحو 3 إلى 4 بليون سنة.[255][256] ولم يحدث أي تغير واضح في مورفولوجيتها أو تنظيمها الخلوي لبضعة بلايين من السنين.[257] نشأت الخلايا حقيقية النواة قبل 1.6 إلى 2.7 بليون سنة. والتغير الكبير التالي في البنى الخلوية حدث حين قامت الخلايا حقيقية النواة بابتلاع البكتيريا في ترابط تعاوني يطلق عليه التعايش الداخلي (بالإنكليزية: endosymbiosis). [258][259] وقد مرت البكتيريا المبتلَعة بتطور مشترك مع الخلايا المضيفة، بحيث تطورت البكتيريا إما إلى متقدرات أو هيدروجينوسومات. [260] وأدى ابتلاع آخر لكائنات حية شبيهة بالزراقم إلى تشكل الصانعات اليخضورية في الطحالب والنبات.[261]
كان تاريخ الحياة يدور حول حقيقيات النواة أحادية الخلايا وبدائيات النواة والعتائق، إلى أن بدأت الكائنات الحية متعددة الخلايا بالظهور قبل 610 مليون سنة في المحيطات خلال الحقبة الإدياكارية (بالإنجليزية).[255][262] لقد حدث تطور تعدد الخلايا في عدة أحداث منفردة في كائنات حية مثل الاسفنجيات، الطحالب البنية، الزراقم، العفن الغروي، والجراثيم المخاطية.[263]
بعد وقت قصير من ظهور الكائنات متعددة الخلايا، ظهر تنوع حيوي بارز عبر 10 ملايين سنة تقريباً في حدث يعرف بانفجار كامبري. وهنا ظهرت في السجل الأحفوري غالبية شعب الحيوانات الحالية بالإضافة إلى بعض السلالات الفريدة التي انقرضت فيما بعد.[264] تم اقتراح مسببات عديدة للانفجار الكامبري، بما فيها تراكم الأكسجين الناتج عن التمثيل الضوئي في الغلاف الجوي.[265] قبل نحو 500 مليون سنة، استعمرت النباتات والفطريات الأرض، وتبعتها بعد قليل المفصليات وحيوانات أخرى.[266] كانت الحشرات ناجحة على وجه الخصوص، وحتى أنها تشكل اليوم الغالبية من أنواع الحيوانات.[267] ظهرت البرمائيات أول مرة قبل حوالي 364 مليون سنة، وتبعتها الحيوانات السلوية البدائية والطيور قبل نحو 155 مليون سنة (وكلاهما من السلالات الشبيهة بالزواحف)، وتلتها الثدييات قبل نحو 129 مليون سنة، وأسرة الإنسانيات قبل نحو 10 ملايين سنة، ومن ثم الإنسان الحديث قبل نحو 250000 سنة.[268][269][270] ولكن رغم تطور هذه الحيوانات الكبيرة، إلا أنَّ الكائنات الحية الأصغر الشبيهة بالتي تطورت في أول هذه السيرورة تبقى ناجحة جداً وهي السائدة على الأرض، إذ أن بدائيات النوى تشكل الغالبية من الأنواع وكذلك الكتلة البيولوجية.
التطبيقات
المفاهيم والنماذج المستعملة في علم الأحياء التطوري، كالاصطفاء الطبيعي، لديها تطبيقات كثيرة.[271] الاصطفاء الاصطناعي هو الانتقاء المتعمد للسمات في التجمعات الأحيائية. وقد استخدم لآلاف من السنين في تدجين النباتات والحيوانات.[272] ومؤخراً، أصبح هذا النوع من الانتقاء جزءاً أساسياً في الهندسة الجينية، حيث تستعمل الواسمات المختارة، مثل مقاومة المضاد الحيوي، للتلاعب بالدنا. وبعد دورات متكررة من التطفر والانتقاء، تطورت البروتينات ذات الخواص القيمة، مثل الإنزيمات المعدلة والأجسام المضادة الجديدة، في سيرورة تسمى التطور الموجه.[273]
إنَّ فهم التغيرات التي حدثت خلال تطور يمكن أن يكشف عن الجينات اللازمة من أجل بناء أعضاء من الجسد، والجينات التي قد تكون متعلقة بالأمراض الوراثية.[274] على سبيل المثال، سمكة الكهف العمياء هي سمكة مهقاء فقدت بصرها خلال التطور. وإنَّ جعل تجمعات مختلفة من هذه الأسماك العمياء تتوالد مع بعضها أنتج نسلاً ذو أعين وظيفية بما أنَّ الطفرات المختلفة حدثت في تجمعات منعزلة تطورت بكهوف مختلفة.[275] وهذا ساعد على تمييز الجينات اللازمة للرؤية والتصبغ.[276] في مجال علم الحاسوب، بدأت عمليات محاكاة التطور باستعمال خوارزميات تطورية وحياة اصطناعية في الستينات، وامتدت فيما بعد لتشمل محاكاة الانتخاب الصناعي.[277] أصبح التطور الاصطناعي طريقة استمثال معترف بها على نطاق واسع بفضل أعمال إنجو ريكينبيرج (بالإنجليزية) في الستينات، حيث استخدم استراتيجيات تطورية لحل مسائل هندسية معقدة.[278] الخوارزميات الوراثية بالتحديد أصبحت مشهورة من خلال كتابات جون هولاند (بالإنجليزية).[279] تضمن التطبيقات العملية أيضاً التطور التلقائي لبرامج الحاسوب.[280] تُستعمل الخوارزميات التطورية الآن لحل مسائل متعددة الأبعاد بشكل أفضل من البرامج التي يبتكرها المصممون البشر، وكذلك تستعمل لتحسين تصاميم الأنظمة.
الاستجابات الثقافية والاجتماعية
في القرن التاسع عشر، وبالتحديد بعد نشر كتاب أصل الأنواع عام 1859،فكرة أنَّ الحياة تطورت مثلت مصدراً ناشطاً للنقاشات العلمية التي تتمحور حول تأثيرات نظرية التطور الفلسفية والاجتماعية والدينية. واليوم أصبح الاصطناع التطوري الحديث مقبولاً لدى الغالبية العظمى من العلماء.[47] ولكن ما زال التطور مفهوماً جدلياً ومحل نزاع لدى بعض المؤمنين بوجود إله.[283]
قامت العديد من الديانات والطوائف بالتوفيق بين معتقداتها والتطور بواسطة مفاهيم مثل التطور الربوبي، فيما هناك خلقيون يؤمنون أن التطور تعارضه أساطير الخلق الموجودة في دياناتهم التي تثير العديد من الاعتراضات على التطور.[141][284][285] وكما أظهرت الاستجابات لنشر كتاب أثارات التاريخ الطبيعي للخلق (بالإنكليزية: Vestiges of the Natural History of Creation) عام 1844، كان أكثر جانب جدلي لعلم الأحياء التطوري هو أنَّ مفهوم تطور الإنسان يقترح أنَّ الإنسان لديه أسلاف مشتركة مع القردة العليا، وأن القوى العقلية والأخلاقية للإنسانية تعود لنفس النوع من المسببات الطبيعية التي شكلت السمات الوراثية عند الحيوانات.[286] وفي بعض الدول، لا سيما الولايات المتحدة، أشعلت التوترات بين العلم والدين الجدل الدائر حول التطور والخلق (بالإنجليزية)، وهو صراع ديني يركز على السياسات والتعليم الحكومي (بالإنجليزية).[287] رغم أن مجالات علمية أخرى تتعارض مع التفسيرات الحرفية للكثير من النصوص الدينية، مثل علم الكون[288] وعلوم الأرض،[289] إلا أن علم الأحياء التطوري يواجه اعتراضات أكثر بكثير من قبل المفسرين الدينيين.
في بعض الدول، يمنع القانون المعلمين من مناقشة الأدلة على التطور أو الاصطناع التطوري الحديث. قوانين بعض الدول الأخرى تنص على أن يعلم فقط علم الأحياء التطوري في المناهج العلمية الملائمة. وعالمياً يعلم التطور في المواضيع العلمية وبقليل من الخلاف، باستثناء بعض المناطق في الولايات المتحدة وعدة دول إسلامية أصولية.
في الولايات المتحدة، تسبب قرار محاكمة القرد عام 1925 في جعل موضوع التطور نادر الظهور بكتب المدرسة الثانوية، ولكن تدريجياً قُدِّم من جديد ليعلم في المدارس بعد نحو جيل، وتمت حمايته قانونياً عام 1968. ومنذ ذلك الحين، تم منع تعليم الخلقية -الاعتقاد الديني المنافس- قانونياً، ولكنه عاد بصورة علم كاذب تحت اسم التصميم الذكي، الذي تم منع تعليمه ثانيةً عام 2005.[290]
في الآونة الأخيرة، أصبح الجدل بارزاً أكثر في الدول الإسلامية.[291] علم الأحياء التطوري مدرج في مناهج المدارس الثانوية في معظم هذه الدول. قامت مؤسسات العلوم في أربع عشرة دولة إسلامية (بما فيها باكستان، إيران، تركيا، إندونيسيا، ومصر) بالتوقيع على بيان صادر عن هيئة الإنترأكاديمي (شبكة عالمية لأكاديميات العلوم)، دعماً لتعليم التطور.[292] أما السعودية والسودان فقد حظرتا تعليم التطور بالمدارس.[293] وقد رُوج للخلقية بشدة في تركيا وبعض المجتمعات المهاجرة في أوروبا الغربية، وبالمقام الأول من قبل هارون يحيى.[294] اعتبر الأوروبيون جدل التطور والخلقية قضية أمريكية.[295] ولكن في السنوات الأخيرة أصبح النزاع قضية في عدة دول ومنها ألمانيا، المملكة المتحدة، إيطاليا، هولندا، بولندا، وصربيا.
أقوال المفكرين المسلمين الموافقة للتطور بمفهومه العام
كان لعلماء المسلمين دور في ملاحظة الطبيعة، ودراسة التاريخ الطبيعي للكائنات. ومن دراساتهم تركوا لنا بعض ملاحظاتهم عن هذا الموضوع. كلمة “التطور” في العنوان لا تشير إلى نظرية التطور الحديثة بالتحديد، ولكن إلى فكرة التطور بشكل عام، وهي فكرة سبقت داروين بمئات وآلاف السنين. وهي النظرة إلى تقارب الكائنات الحية وتشابهها، وإمكانية ترتيبها من الأكثر بساطة الى الأكثر تعقيداً، وإعتبارات هذا التقارب وما قد يستنتج منه.
ابن خلدون
يذكر ابن خلدون في كتاب “المقدمة” عن إتصال الكائنات الحية ببعضها: “إنّا نشاهد هذا العالم بما فيه من المخلوقات كلها على هيئة من الترتيب والإحكام، وربط الأسباب بالمسببات واتصال الأكوان بالأكوان، واستحالة بعض الموجودات إلى بعض… انظر إلى عالم التكوين كيف ابتدأ من المعادن ثم النبات ثم الحيوان على هيئة بديعة من التدرج: آخر أفق المعادن متصل بأول أفق النبات مثل الحشائش وما لا بذر له، وآخر أفق النبات مثل النخيل والكرم متصل بأول أفق الحيوان مثل الحلزون والصدف ولم يوجد لهما إلا قوة اللمس فقط. ومعنى الاتصال في هذه المكونات أن آخر أفق منها مستعد بالاستعداد القريب لأن يصير أول أفق الذي بعده. واتسع عالم الحيوان وتعددت أنواعه وانتهى في تدرج التكوين إلى الإنسان صاحب الفكر والروية، ترتفع إليه من عالم القردة الذي اجتمع فيه الحس والإدراك ولم ينته إلى الروية والفكر بالفعل، وكان ذلك أول أفق الإنسان بعده، وهذا غاية شهودنا.”
كما ينفي في نفس الكتاب نسبة أحد الصفات إلى أسباب غيبية غامضة ويدل على الأسباب الطبيعية التي تفسر هذه الظاهرة:
“وقد توهم بعض النسابين ممن لا علم لديه بطبائع الكائنات أن السودان وهم ولد حام بن نوح، اختصوا بلون السّواد لدعوة كانت عليه من أبيه ظهر أثرها في لونه وفيما جعل الله الرّق في عقبه، وينقلون في ذلك خرافات من القصص، ودعاء نوح على ابنه حام قد وقع في التـوراة وليس فيه ذكر السّواد ، وإنما دعا عليه بـأن يكون ولده عبيدا لولد أخوته لا غير . وفي القول بنسبة السودان إلى حام. غفلة عن طبيعة الحر والبرد وأثرهما في الهواء وبما يتكون فيه من الحيوانات ، وذلك أن هذا اللون شمل أهل الإقليم الأول والثاني من مزاج هوائهم للحرارة المتضاعفة في الجنوب ، فإن الشّمس تسامت رؤسهم مرّتين في كل سنة قريبة إحداهما من الأخرى فتطول المساتمة عامة الفصول ، فيكثر الضوء لأجلها ويلح القيظ الشديد وتسود جلودهم لإفراط الحر.”
إبن مسكويه
يقول إبن مسكوية في كتابه “تهذيب الأخلاق وتطهير الأعراق”: “أن الموجودات مراتب، وكلها سلسلة متصلة… وكل نوع من الموجودات يبدأ بالبساطة ثم لا يزال يترقى ويتعقد حتى يبلغ أفق النوع الذي يليه، فالنبات في أفق الجماد، ثم يترقى حتى يبلغ أعلى درجة، فإذا زاد عليها قبل صورة الحيوان. وكذلك الحيوان يبدأ بسيطاً ثم يترقى حتى يصل إلى مرتبة قريبة من الإنسان.”
ويضيف عن تدرج الحيوان إلى ما يقترب من صورة الإنسان قائلاً:
“هو الذي يحاكي الإنسان من تلقاء نفسه ويتشبه به من غير تعليم كالقردة وما أشبهها، ويبلغ من ذكائها أن تستكفي في التأدب بأن تري الإنسان يعمل عملاً فتعمل مثله من غير أن تحوج الإنسان إلى تعب بها ورياضة لها. وهذه غاية أفق الحيوان التي إن تجاوزها وقبل زيادة يسيرة، خرج بها عن أفقه وصار في أفق الإنسان الذي يقبل العقل والتمييز والنطق والآلات التي يستعملها والصور التي تلائمها.” [١]
الجاحظ
كان الجاحظ أول أحيائي وفيلسوف مسلم يتكهن بالتطور تفصيليا في القرن التاسع للميلاد. في كتابه الحيوان، أخذ بعين الاعتبار التأثيرات البيئية على فرص الحيوان للبقاء، ووصف الصراع من أجل البقاء، وكتب الجاحظ أيضا عن سلاسل الغذاء. يعتبر تكهن الجاحظ عن تأثير البيئة على الحيوانات نوع مبكر من نظريات التطور. ووصف الصراع من أجل البقاء حيث توقع فيه الاصطفاء الطبيعي.”
القزويني
أما القزويني، وهو من مشاهير العلوم الطبيعية المسلمين فيذكر في كتابه “عجائب المخلوقات وغرائب الموجودات”: “إن أول مراتب هذه الكائنات تراب وآخرها نفس ملكية طاهرة، فالمعادن متصلة أولها بالتراب أو الماء وآخرها بالنبات. والنبات متصل أوله بالمعادن وآخره بالحيوان، والحيوان متصل أوله بالنبات وآخره بالإنسان، والنفوس الإنسانية متصلة أولها بالحيوان وآخرها بالنفوس الفلكية”
خالص جلبي
يقول المفكر الإسلامي خالص جلبي في مقاله “متى بدأت رحلة الإنسان على الأرض؟“: “عندما كتب المؤرخون عن تاريخ الإنسان على الأرض كانت الرؤية التقليدية أنه هبط من جنة عدن إلى أرض الشقاء. وكلمة (اهبطوا) أوحت إلى البعض أنه (أنزل) من كوكب آخر. وأنه عجن دفعة واحدة. وأن لا علاقة له ببقية الكائنات.[...] وما يقوله العلم ان آدم عاش في أفريقيا وأنه لم يكن شكلاً واحداً بل زاد عن عشرة أشكال. وأنه كان من طين لازب من الأرض. وأنه وبقية الكائنات تتصل بشجرة الخليقة فلم يهبط آدم من السماء بل نبت من الأرض والله أنبتكم نباتا.[...] وكان (ابن خلدون) و(ابن مسكويه) من أوائل من أشار إلى اتصال كل أفق من المخلوقات بالأفق الذي بعده كما بين الحلزون والنخيل وأن هناك نوعا من التحول (العجيب) من آخر أفق من الكائنات إلى أول الأفق الذي بعده. ومع أن (تشارلز دارون) طرح منذ عام 1859 كتابه عن (أصل المخلوقات) فنحن ما زلنا في الشرق لا نتحدث في هذا الموضوع خوفا من العوام. كما حصل عام 1854م في زمن الشريف (عبد المطلب) في مكة حينما جاءه أمر من الوالي العثماني بأن السلطان تحت ضغط بريطانيا يريد إنهاء بيع الرقيق في الأسواق. (فلما استدعى الوالي دلالي الأرقاء وأبلغهم بالخبر هاج الناس وتنادوا بالجهاد.. وأنه مخالف للشرع واشتبكوا مع الحامية التركية ووقع العديد من القتلى) على ما ذكره الوردي في موسوعته عن تاريخ العراق الحديث.”
فالفكرة واضحة لمن يتأمل الطبيعة ويدرسها بعمق، ولا يوجد تعارض مبدئي بينها وبين الاسلام حسب فهم عدد من مفكري الاسلام وعلماء الطبيعة المسلمين على مر العصور. هذا كله اشارة الى قبولهم لاتصال افق الكائنات الحية ببعضه. وأيضاً على أفضلية التفسير الطبيعي العلمي للظواهر الطبيعة.
المصدر
الموسوعة الحرة ويكيبديا
+
مدونة جهاد
+
يوتيوب















 رد مع اقتباس
رد مع اقتباس